Microfossiel
Dit artikel is in bewerking voor de Schrijfwedstrijd.
Wil je een wijziging in dit artikel doorvoeren, dan graag eerst de beoogde wijziging melden op de overlegpagina. Voor meer informatie hierover zie hier.

Microfossielen zijn microscopisch kleine fossiele overblijfselen van organismen die tussen de 0,001 mm en 1 mm groot zijn. Ze kunnen geobserveerd worden met een lichtmicroscoop of een elektronenmicroscoop. Macrofossielen kunnen met het blote oog of met een vergrootglas bekeken worden.
Microfossielen kunnen complete organismen zijn zoals prokaryoten met als voorbeeld de cyanobacteriën. Het kunnen ook eencellige eukaryoten zijn zoals de foraminifera of delen daarvan zoals de kalkplaatjes of coccolieten van de coccolithoforen. Het kan ook gaan om delen van grotere dieren of planten zoals kleine tanden of sporen van schimmels en planten.
Microfossielen met een exoskelet zoals de coccolithoforen en de foraminifera, dragen als zodanig bij aan de sedimenten en kunnen kalkstenen rotsen vormen, zoals de krijtrotsen van Dover. Andere microfossielen, die geen exoskelet bezitten, kunnen ook een soort afzettingen vormen, zoals de stromatolieten. Deze bestaan uit biofilmen of bacteriële matten gevormd door cyanobacteriën. Behalve voor de micropaleontologie, zijn de microfossielen ook zeer belangrijk voor de biostratigrafie, waarbij gesteenten ten opzichte van elkaar gedateerd kunnen worden. Ze zijn ook van groot belang voor de bestudering van het paleoklimaat.
Microfossielenbestand
Het microfossielenbestand loopt vanaf het begin van het leven in het Precambrium, van 3500 miljoen jaar geleden (3,5 Ga) tot aan het heden, in het Kwartair. Microfossielen komen het meest voor in afzettingen in oceaan- en zeebodems, maar ze komen ook voor in brak water, zoet water en in sedimentaire afzettingen op het land. De meest voorkomende vormen van microfossielen zijn exoskeletten van protisten en cysten van microben, samen met stuifmeel en sporen van de vaatplanten. Eerst zijn de meest voorkomende fossielen uit de jongere perioden gevonden, maar uiteindelijk zijn in meer dan 3 miljard jaar oude gesteenten verschillende microfossielen van ongeïdentificeerde prokaryoten gevonden. Door een beter begrip van soortconcepten, van de verspreidingsgebieden en door de volledigheid van het microfossielenbestand kunnen nu het hele Fanerozoïcum vanaf 451 miljoen jaar (451Ma) geleden tot heden en delen van het Proterozoïcum, van 2,5 Ga geleden tot aan het Fanerozoïcum, worden gedateerd met behulp van microfossielen. Sommige microscopische groepen, zoals de cyanobacteriën, zijn met hun opkomst aan het begin van het Proterozoicum (2,5 Ga geleden), verantwoordelijk voor de zuurstofrijke atmosfeer die ontstond en als gevolg waarvan er een zuurstofcrisis begon. Deze micro-organismen vormden als eerste een atmosfeer met zuurstof waarmee de Aarde een voor ons bewoonbare planeet is geworden.[1]
Biostratigrafie


Gidsfossielen zijn fossielen die gebruikt worden voor het dateren van afzettingen, ook wel sedimenten genoemd. Deze vorm van datering met gebruik van microfossielen is de biostratigrafie, die aan de hand van fossielen de geologische tijd bepaalt en zo biologie en geologie samenbrengt. Biostratigrafie onderzoekt de relatie tussen soorten micro-organismen, hun omgevingen en hun evolutie door de tijd heen, uitgedrukt in exoskeletten bewaard als fossielen. Dit onderzoek ontwikkelde zich als een discipline die essentieel was voor de groei van de historische wetenschap en was gebaseerd op drie fundamentele aannames:
- vaststellen van opeenvolgende verzamelingen van fossielen in opeenvolgende lagen;
- testen en bevestigen van die opeenvolging in andere plaatsen en andere regio's;
- komen de verzamelingen fossielen overeen dan hebben ze een vergelijkbare geologische leeftijd.
Microfossielen als acritarchen, chitinozoën, tanden van conodonten, cysten van dinoflagellaten, ostracoden, stuifmeel, sporen en foraminiferen behoren tot de vele soorten die zijn geïdentificeerd als gidsfossielen die veel worden gebruikt in biostratigrafie. De absolute leeftijd van de lagen wordt echter bepaald met onderzoek naar de aanwezige radioactieve isotopen, terwijl de namen van de stratigrafische eenheden, de biozones, worden onderscheiden al naar gelang de aanwezige fossielen. Microfossielen zijn bij uitstek geschikt voor biostratigrafische analyse omdat ze bijzonder overvloedig aanwezig kunnen zijn in gesteenten en omdat ze kunnen worden gewonnen met relatief eenvoudige methoden van massale verwerking. Veel groepen zijn geografisch verspreid (bijvoorbeeld plankton, sporen in de lucht en stuifmeel). Wanneer ze snel evolueren wordt de stratigrafische resolutie hoger.
Paleoklimatologie
Microfossielen zijn ook goede indicators voor het klimaat uit het verleden en vormen studieobjecten voor de paleoklimatologie. Foraminifera bijvoorbeeld komen overal voor, in zee en aan het land en van de polen tot de tropen. De isotopen van hun kalkschaaltjes (CaCO3) registreren temperatuur en chemische samenstelling van de oceanen en vormen daarmee indicatoren voor het klimaat van het Mesozoïcum (252 Ma geleden) tot het Kwartair, het huidige geologische tijdperk.
De draaiing van de opeenvolgende kamers in het schelpje van sommige forminifera, kan aangeven wat de temperatuur van het water geweest moet zijn. Dit is het geval bij Neogloboquadrina pachyderma (eerder Globigerina pachyderma genaamd) waarbij subpolaire en polaire populaties werden onderscheiden door een overheersende links- of rechtsom draaiende spiraal. De verspreiding van deze twee fenotypen correleert sterk met het circulatiepatroon aan het oppervlaktewater. De een geeft de voorkeur aan koud water en de ander aan warm water. De geschiedenis van de Kwartaire temperatuurschommelingen van de oceaan kan daarom worden bepaald aan de hand van de verspreiding van planktonforaminiferen die bewaard zijn gebleven in diepzeekernen. Deze linker en rechter spiraalvormen werden lange tijd als twee verschillende fenotypen gezien, zogenaamde ecofenotypen. Uit moleculair genetisch onderzoek is echter gebleken dat het verschillende soorten zijn. N. pachyderma is de linker kronkelende vorm, terwijl de rechter kronkelende vorm, die warm water prefereert, N. incompta wordt genoemd.[2][3]
Biomineralisatie

De overgang van het Ediacarium naar het Cambrium, tussen 550 en 530Ma geleden, documenteert een belangrijke fase in de diversificatie van dieren. Het Cambrium wordt gemarkeerd door de plotselinge ‘explosie’ van nieuwe organismen die wel of niet gebiomineraliseerd waren. In het totale fossielenbestand uit die tijd verschijnen Cloudina-achtige en andere buisvormige organismen met een schaal, gevolgd door kleine koolstofhoudende fossielen zonder biomineralisatie en zeer gevarieerde kleine schelpachtige fossielen. In 2024 zijn in de Bocaina formatie in Brazilië fossiele vondsten van dieren gedaan, die lijken op Cloudina uit het Ediacarium en die deze overgangsfase meer dan 20 miljoen jaar terug zouden brengen.
Een belangrijke stap in deze overgang was biomineralisatie. De microfossielen uit het Ediacarium zijn hoofdzakelijk acritarchen (eukaryotische eencelligen zonder extern skelet) die reeds aanwezig waren ook eerder in het Proterozoïcum, evenals de stromatolieten en bacteriën. Het ontstaan van het exoskelet is een nieuwe en belangrijke ontwikkeling in de evolutie en in de geschiedenis van de Aarde. Het skelet heeft de dieren mogelijk beschermd tegen predatoren. De trechter-in-trechtervorm van het externe skelet van Cloudina bestaat uit calciet. Het diertje is meercellig en het is meerdere millimeters groot. Het is eigenlijk geen microfossiel, maar staat symbool voor het begin van de biomineralisatie.[4]
Indeling
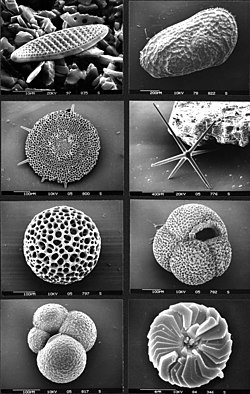
Microfossielen kunnen worden onderverdeeld in twee hoofdgroepen. Deze groepen zijn niet taxonomisch maar maken onderscheid in de samenstelling van het exoskelet en de fossiele resten: de minerale en organische microfossielen die bestudeerd worden door twee verschillende studiegebieden, respectievelijk de micropaleontologie en de palynologie:
Micropaleontologie: Dit vak bestudeert microfossielen van organismen of delen daarvan die ook gedurende hun leven minerale structuren bezitten:
- Kalkhoudende microfossielen: coccolithoforen, foraminifera, mosselkreeftjes (ostracoda) en kalkhoudende cysten van dinoflagellaten.
- Siliciumhoudende microfossielen: diatomeeën, fytolieten, stralendiertjes (radiolaria) en skeletnaalden van bijvoorbeeld sponzen.
- Fosfaathoudende microfossielen: scolecodonten (stukjes kaak), conodonten (kleine stukjes van monddelen), graten en tanden van haaien en andere overblijfselen van vissen.
Palynologie: De organische microfossielen zijn onderwerp van studie in de palynologie en worden palynomorfen genoemd. Dit zijn microfossielen van organismen of delen ervan die gedurende hun leven een externe organische wand hebben, zonder mineralen. Tot deze groep behoren de sporen, schimmels, acritarchen, stuifmeel, chitinozoa, scolecodonts (stukjes kaak), cysten van dinoflagellaten.
Minerale microfossielen
Kalkhoudend
Kalkhoudend kan verwijzen naar calciet of aragoniet, wat beide verschillende kristalstructuren van calciumcarbonaat (CaCO3) zijn, die de plaatjes, schelpjes of schaaltjes van microfossielen vormen en ook de afzettingen daarvan. Enkele van de belangrijkste microfossielen die kalkskeletjes produceren zijn coccolithoforen en foraminifera.
Coccolithoforen

Bij de eencellige microalgen zoals de coccolithoforen bijvoorbeeld, zijn het de coccolieten of kalkskeletjes van calciumcarbonaat die als fossiel overblijven en kalksteen vormen. Dit gebeurde voornamelijke gedurende het laat-Krijt en wordt ook wel het krijtgesteente genoemd. Gedurende deze periode van enorme diversificatie van de soorten zijn de coccolithoforen verspreid over grote gebieden. Voorbeelden van omvangrijk krijtgesteente zijn de kliffen van Dover en van Calais. Het is nog niet duidelijk waar de coccolieten voor dienen. Ze kunnen dienen als bescherming tegen zonlicht of juist om het zonlicht beter te concentreren, als depot voor de giftige calciumionen of als ballast om de cel beter te stabiliseren. De meeste coccolithoforen zijn uitgestorven tussen het Krijt en het Paleogeen en hun habitats werden daarna overgenomen door de diatomeeën. Nadien zijn ze de tropicale en gematigde wateren gaan domineren, maar ze zijn nooit meer zo gevarieerd geweest als in het Mesozoïcum.
Foraminifera

Foraminifera zijn eencellige eukaryoten die, net zoals de coccolithoforen, een schelpje of schaal bezitten van calciet. Deze schaaltjes kunnen een of meerdere kamers vormen die verbonden zijn door een of meerdere openingen (foramina). De schaaltjes kunnen ook gemaakt zijn van organisch materiaal als tectine, een chitine-achtige substantie of andere mineralen zoals aragoniet of silicium. Ook kan de schaal gevormd worden door aan de tectine geplakte zandkorreltjes, maar dan hebben ze als microfossiel weinig waarde. Ze komen voor sinds het vroege Cambrium (541 Ma geleden) en bereikten hun hoogtepunt in het Cenozoïcum tot aan vandaag. Hun schaaltjes zijn overvloedig aanwezig en kunnen al naar gelang de soort, de meest verschillende vormen hebben. Ze maken voor 55% deel uit van de Arctische biomassa en voor meer dan 90% deel uit van de diepzee-biomassa. Omdat ze overvloedig, gevarieerd en makkelijk te bestuderen zijn, vormen ze belangrijke biostratigrafische indicatoren voor het Laat-Paleozoïcum, Mesozoïcum en Cenozoïcum (vanaf 450 Ma geleden tot nu).
Silicium
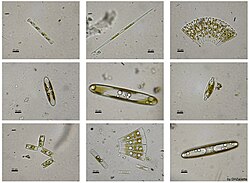
Ook diatomeeën zijn eencellige algen, die evenals sommige cyanobacteriën koloniën kunnen vormen, meestal ketens. De cellen bezitten een exoskelet van siliciumdioxyde (SiO2). Het exoskelet wordt frustule genoemd wat verwijst naar een doosje met een dekseltje: een hypotheca en epitheca die kleine poriën of punctae bevatten met zeer uiteenlopende patronen. Deze patronen bepalen de classificatie van de diatomeeën die ook afhangt van andere morfologische aspecten van het exoskelet. Over het algemeen zijn ze tussen de 1 en 100 μm groot, maar ze kunnen ook lange ketens van meerder frustules vormen. Ze zijn autotroof en staan aan de basis van de voedselketen. Ze leven zowel in zoet water als in zeewater maar ook in de bodem of op rotsen. Ze hebben licht nodig en daarom blijven ze binnen de fotische zone (<200m diepte). In bepaalde omstandigheden, wanneer er veel voedsel en mineralen beschikbaar zijn, kunnen er wel 1 miljard cellen per m3 zeewater aanwezig zijn. Elk soort diatomee geeft de voorkeur aan een bepaald bereik in temperatuur, zoutgehalte, zuurtegraad, zuurstof en mineralen. Slechts 5% van de levende diatomeeën aan het oppervlak bereikt de bodem als frustules of statosporen (cysten met silicium wand en bijbehorende frustules). De sedimenten zijn daarom moeilijk te interpreteren ook omdat diatomeeën zich ver kunnen verplaatsen alvorens ze de zeebodem bereiken. Denk bijvoorbeeld aan de zoetwater diatomeeën die van het land de zee inwaaien. Het fossielenbestand van diatomeeën is slechts gedeeltelijk bekend omdat ze makkelijk oplossen en vanwege tafonomische effecten. De eerste duidelijke frustules dateren van het Vroeg-Jura (145 Ma geleden).
Organische microfossielen
Palynomorfen

Palynomorfen zijn microfossielen van organisch materiaal. Dat kunnen uiteenlopende plantenresten, dierenresten of resten van eencelligen zijn. Veel gebruikte microscopisch kleine fossiele resten in palynomorfen zijn dinoflagellaten, acritarchen, sporen, pollen, schimmels, scolecodonten, organen van wormen, van borstelwormen of van geleedpotigen (zoals monddeeltjes van insecten), chitinozoa en foraminifera.
De oudste sporen

Sporen hebben een organische taaie wand die resistent is tegen temperatuur, mechanische druk en micro-organismen. Deze resistentie maakt het mogelijk reproductieve cellen le laten overleven in ongunstige omstandigheden. Ze maken deel uit van de levenscyclus van schimmels, algen, mossen en varens. De oudste sporen dateren van 470 - 460 Ma geleden in de periode Ordovicium. In deze periode hebben de landplanten zich ontwikkeld. Er zijn dan voornamelijk twee soorten plantensporen: de cryptosporen en de trilete sporen. De eerste zijn onderdeel van de levenscyclus van planten die geleken moeten hebben op Bryophyta (mossen) terwijl de trilete sporen behoorden tot de voorlopers van de vaatplanten, de Tracheophyta.[5] Deze laatste sporen ondergingen meiose en vormden een tetrade van gameten. Deze oude sporen behoorden tot een levenscyclus die geërfd was van de groene algen en ze brachten de eerste landplanten voort die inmiddels uitgestorven zijn.
Ontdekkingen van de oudste microfossielen
Fossielen uit het Cambrium en daarna, zijn eerder en makkelijker ontdekt dan die van het voorgaande Precambrium en ze bevestigden de evolutietheorie van Charles Darwin. Er is lang getwijfeld of er leven geweest is in de allervroegste eonen. Darwin was verbaasd over de afwezigheid van fossielen uit het Precambrium wat een sterk argument geweest zou zijn tegen zijn evolutietheorie. In 1883 ontdekte Charles Doolittle Walcott voor het eerst stromatolieten uit het Precambrium die hij Cryptozoon noemde. Stromatolieten zijn gesteenten die ontstaan uit microben die matten vormen. In 1899 ontdekte hij millimeters grote donkere ronde plaatjes, die hij Chuaria noemde. Daar werd toen van gedacht dat het geplette schelpen waren. Pas later bleek dat het om echte eencellige algen ging, de eerste gevonden fossiele cellen uit het Precambrium.[6] Deze eukaryotische cellen zijn te groot om tot de microfossielen gerekend te worden, maar ze vormen de eerste gevonden fossiele cellen uit het Precambrium.
Twijfel en ongeloof

Het ongeloof en sceptiscime ook ten aanzien van de Cryptozoon was groot en verder onderzoek in dit veld stagneerde totdat Stanley A. Tyler in 1953 de Gunflint Chert ontdekte, een geologische formatie gelegen tussen Ontario (Canada) en Minnesota (VS) daterend uit halverwege het Precambrium, rond de 1,9 Ga (1,9 miljard jaar). Met gebruik van de microscoop trof hij daar in koolstofhoudende vuurstenen bolvormige en draadvormige microfossielen aan.[8]
Er ontstond opnieuw twijfel over de vraag waarom er uitsluitend in de Gunflint Chert microfossielen gevonden waren en ook alleen uit die tijd, 1,9 Ga geleden. Vervolgens werden echter in 1965 draadvormige microfossielen gevonden in Precambrische stromatolitische zwarte vuursteen uit Bitter Springs Chert, Centraal Australië die dateerden van 1 Ga geleden. Deze fossielen waren beter bewaard dan die van Gunflint Chert.[6]
Uiteindelijk werd in 1992 een verzameling gepermineraliseerde draadvormige microfossielen en bijkomend organisch materiaal in de 3,5 Ga oude Apex Chert in het noordwesten van Australië ontdekt en onderzocht. Deze microfossielen worden algemeen beschouwd als een van de oudste getuigenissen van leven: ze leefden 1 Ga na het ontstaan van het zonnestelsel.[9] Er zijn oudere vondsten gedaan van 3,95 Ga, maar daar is nog weinig zekerheid over.[10][11] Vrijwel alle gevonden microfossielen uit het Precambrium blijven een bron van twijfel. Wetenschappers worden het er niet over eens.
Het is namelijk aangetoond dat micellen en filamenten die in het laboratorium met vetzuren gevormd zijn, in een medium met verschillende zouten, gelijkenis vertonen met microfossielen wanneer men ze observeert met de confocale en elektronenmicroscoop. De micellen en filamenten zouden, net als de microfossielen uit het Precambrium, voorlopers kunnen zijn van het membraan dat de cel nog moet omgeven. De microfossielen zouden dus, net als de micellen in het laboratorium, abiotisch (zonder leven) geweest kunnen zijn. [12]
Precambrium


Reeds uit het begin van het Precambrium, zo'n 3,5 miljard jaar geleden, zijn er microfossielen gevonden. De microfossielen uit die tijd hebben de meeste kenmerken voor identificatie verloren, maar bezitten regelmatige structuren, zoals filamenten. Ze blijven onderwerp van debat. Een recentere vorm die wel geïndentificieerd kan worden is die van Gunflintia, een uitgestorven cyanobacterie, die een van de eerste zuurstofproducerende organismen geweest is. Het fossiel is het meest voorkomende in de Gunflint Chert. Ook in het huidige tijdperk kunnen cyanobacteriën zich organiseren in filamenten of draden.
Het eon Proterozoïcum, de tijd waarin de Gunflint Chert gevormd werd, begint met de zuurstofcrisis, waarin de atmosfeer relatief plotseling met zuurstof verrijkt werd door de cyanobacteriën. Dit eon kent ook vondsten van Acritarchen die gedefinieerd worden als microfossielen van onbekende oorsprong, maar die in de meeste gevallen eencellige eukaryoten blijken te zijn.[14] De meeste werden gevonden in gesteenten van 1,8 Ga geleden, maar vondsten in 2,1 Ga oude gesteenten zijn gemeld. Af en toe wordt een gevonden specimen geïdentificeerd als een eencellige alg of spore en verdwijnt daarmee van de lijst gevonden Acritarchen. Er bestaat nog veel onzekerheid over de identiteit van de Acritarchen.
Ediacarium
Aan het eind van het Proterozoïcum, tijdens het Ediacarium, ontstaan er meercellige macroscopische dieren. Het oudste overtuigende fossiele bewijsmateriaal voor meercellige dieren is ongeveer 600 miljoen jaar oud. Volgens sommige onderzoekers is dit echter veel jonger dan wat de studie van het rRNA (ribosomaal RNA) van levende organismen suggereert. De dieren zouden dus al langer hebben moeten bestaan. De sequenties van genen evolueren met een zekere regelmaat waardoor verschillen kunnen worden gebruikt als moleculaire klokken. Zo kan worden berekend dat de afstammingslijnen van ongewervelde dieren ongeveer 1,2 Ga geleden uit elkaar zijn begonnen te lopen. Dit zou betekenen dat de eerste dieren ontbreken in het fossielenbestand, ze zouden niet gefossiliseerd zijn. Dat zou kunnen komen doordat ze bijvoorbeeld in de waterkolom hebben geleefd of als microscopische organismen in het sediment leefden. De zogenaamde Cambrische explosie rond 541 miljoen jaar geleden zou dan voor een groot deel te maken hebben met de groei van (exo)skeletten, waar vervolgens fossielen van overgebleven zijn.
Cambrium
Metazoa (meercellige dieren) scheiden sinds het begin van het Cambrium en ook eerder, zoals Cloudina liet zien, kalkhoudende skeletten af, maar bij de protozoa (eencellige dieren) gaat het anders. Hoewel benthische foraminiferen met organische wanden en samengeplakte omhulsels in het vroege Cambrium verschenen, kwamen vormen met CaCO3-schalen pas in het Devoon tot het Carboon voor en sindsdien zijn ze geleidelijk uitgebreid. Deze vertraging in de biomineralisatie met calciet (CaCO3) van protozoa kan worden verklaard door de hoge oppervlakte-volumeverhouding van een enkele cel, waardoor de aanleg van een schaal of schelp relatief veel energie en materiaal kost. Bovendien zijn de protozoa gevoeliger voor waterchemie, zoals de zuurtegraad, dan complexere metazoa.
Bronnen
- Howard A. Armstrong, Martin D. Brasier (2005) Microfossils. Blackwell publishing ISBN 0-632-05279-1
- Brian McGowran (2005) Biostratigraphy, Microfossil and Geological Time Cambridge University Press ISBN 978-0-521-83750-7
Referenties
- ↑ (en) Knoll, Andrew H., Nowak, Martin A. (5 mei 2017). The timetable of evolution. Science Advances 3 (5). ISSN:2375-2548. PMID: 28560344. PMC: PMC5435417. DOI:10.1126/sciadv.1603076.
- ↑ pforams@mikrotax - Neogloboquadrina pachyderma. www.mikrotax.org. Geraadpleegd op 8 augustus 2024.
- ↑ (en) WoRMS - World Register of Marine Species - Neogloboquadrina incompta (Cifelli, 1961). www.marinespecies.org. Geraadpleegd op 8 augustus 2024.
- ↑ a b (en) Morais, Luana, Freitas, Bernardo T., Fairchild, Thomas Rich, Clavijo Arcos, Rolando Esteban, Guillong, Marcel (28 juni 2024). Dawn of diverse shelled and carbonaceous animal microfossils at ~ 571 Ma. Scientific Reports 14 (1): 14916. ISSN:2045-2322. DOI:10.1038/s41598-024-65671-4.
- ↑ a b (en) Rubinstein, Claudia V., Vajda, Vivi (3 juli 2019). Baltica cradle of early land plants? Oldest record of trilete spores and diverse cryptospore assemblages; evidence from Ordovician successions of Sweden. GFF 141 (3): 181–190. ISSN:1103-5897. DOI:10.1080/11035897.2019.1636860.
- ↑ a b (en) Schopf, J. William (20 juni 2000). Solution to Darwin's dilemma: Discovery of the missing Precambrian record of life. Proceedings of the National Academy of Sciences 97 (13): 6947–6953. ISSN:0027-8424. PMID: 10860955. PMC: PMC34368. DOI:10.1073/pnas.97.13.6947.
- ↑ (en) Schirrmeister, Bettina E., Sanchez-Baracaldo, Patricia, Wacey, David (2016-07). Cyanobacterial evolution during the Precambrian. International Journal of Astrobiology 15 (3): 187–204. ISSN:1473-5504. DOI:10.1017/S1473550415000579.
- ↑ (en) Barghoorn, Elso S., Tyler, Stanley A. (5 februari 1965). Microorganisms from the Gunflint Chert: These structurally preserved Precambrian fossils from Ontario are the most ancient organisms known.. Science 147 (3658): 563–575. ISSN:0036-8075. DOI:10.1126/science.147.3658.563.
- ↑ Schopf, J. William, Kudryavtsev, Anatoliy B. (1 november 2012). Biogenicity of Earth's earliest fossils: A resolution of the controversy. Gondwana Research 22 (3): 761–771. ISSN:1342-937X. DOI:10.1016/j.gr.2012.07.003.
- ↑ (en) Tashiro, Takayuki, Ishida, Akizumi, Hori, Masako, Igisu, Motoko, Koike, Mizuho (2017-09). Early trace of life from 3.95 Ga sedimentary rocks in Labrador, Canada. Nature 549 (7673): 516–518. ISSN:1476-4687. DOI:10.1038/nature24019.
- ↑ (en) Magazine, Smithsonian, This May Be the Oldest Traces of Life Yet Found. Smithsonian Magazine. Geraadpleegd op 4 augustus 2024.
- ↑ (en) Jordan, Seán F., van Zuilen, Mark A., Rouillard, Joti, Martins, Zita, Lane, Nick (10 mei 2024). Prebiotic membrane structures mimic the morphology of alleged early traces of life on Earth. Communications Earth & Environment 5 (1): 1–13. ISSN:2662-4435. DOI:10.1038/s43247-024-01372-0.
- ↑ (en) Maldanis, L., Hickman-Lewis, K., Verezhak, M., Gueriau, P., Guizar-Sicairos, M. (18 mei 2020). Nanoscale 3D quantitative imaging of 1.88 Ga Gunflint microfossils reveals novel insights into taphonomic and biogenic characters. Scientific Reports 10 (1): 8163. ISSN:2045-2322. DOI:10.1038/s41598-020-65176-w.
- ↑ (en) Jain, Sreepat (12 december 2019). Fundamentals of Invertebrate Palaeontology: Microfossils. Springer Nature. ISBN 978-81-322-3962-8.










